Ваша моцарелла не тянется или горит? Секрет в науке. От процесса «паста филата» до решающей роли созревания...
Поиск в блоге
Категории блога
Последние посты

Секрет идеальной тянущейся моцареллы: научное руководство по приготовлению пиццы
Читать далее

Французский морковный салат | Аутентичный рецепт и профессиональные советы Читать далее
Французский морковный салат (Carottes Râpées) - это полезная и вкусная закуска, которая готовится менее чем за 15...

Человек и времена года: Влияние смены сезонов на здоровье, психику и образ жизни Читать далее
Знаете ли вы, что смена сезонов влияет на ваше здоровье, настроение и даже гены? Узнайте, как жить в гармонии с...

Эффект Даннинга-Крюгера: когда невежество притворяется уверенностью Читать далее
Что такое эффект Даннинга-Крюгера? Узнайте, как этот феномен влияет на нашу жизнь, и научитесь избегать его ловушек...

Подсолнечный мед: свойства, польза и полное руководство от эксперта Читать далее
Узнайте о подсолнечном меде, его лечебных и биохимических свойствах, истории и способах определения чистого меда.
Популярные посты

Мёд и диабет: от молекулярных механизмов до клинических протоколов применения
Комплексный научный обзор роли мёда в управлении диабетом. От влияния на клеточные пути и клинических данных до...
Читать далее
Медовый хаб: ведущий поставщик натуральных продуктов пчеловодства в Иране
Медовый хаб - это ведущий поставщик натуральных продуктов пчеловодства в Иране. Мы предлагаем широкий ассортимент...
Читать далее
Удивительный состав сырого меда | Почему важны ферменты меда?
Сырой мед - это сочетание удивительных питательных веществ, в том числе нескольких ферментов. Эти ферменты играют...
Читать далее
Все, что вам нужно знать о меде
Мед - это натуральный и полезный продукт, который имеет много пользы для здоровья. В этом посте мы дадим вам полную и...
Читать далее")
Маточное молочко: польза и вред, как принимать — полное научное руководство (2025)
Откройте для себя мощный суперфуд, создающий пчелиных маток. Полное руководство по его пользе для омоложения,...
Читать далееПопулярные сообщения
Секрет идеальной тянущейся моцареллы: научное руководство по приготовлению пиццы
Ваша моцарелла не тянется или горит? Секрет в науке. От процесса «паста филата» до решающей роли созревания...
Читать далее Французский морковный салат | Аутентичный рецепт и профессиональные советы
Французский морковный салат (Carottes Râpées) - это полезная и вкусная закуска, которая готовится менее чем за 15...
Читать далее Человек и времена года: Влияние смены сезонов на здоровье, психику и образ жизни
Знаете ли вы, что смена сезонов влияет на ваше здоровье, настроение и даже гены? Узнайте, как жить в гармонии с...
Читать далее Эффект Даннинга-Крюгера: когда невежество притворяется уверенностью
Что такое эффект Даннинга-Крюгера? Узнайте, как этот феномен влияет на нашу жизнь, и научитесь избегать его ловушек...
Читать далее Подсолнечный мед: свойства, польза и полное руководство от эксперта
Узнайте о подсолнечном меде, его лечебных и биохимических свойствах, истории и способах определения чистого меда.
Читать далее
 شکل 1- ملکهها دو نوع فرمون دارند: 1- فرمون آغازگر 2- فرمون آزادکننده
شکل 1- ملکهها دو نوع فرمون دارند: 1- فرمون آغازگر 2- فرمون آزادکننده شکل 2- سیگنال های ملکه و انواع فرمون ها برای مدیریت کندو
شکل 2- سیگنال های ملکه و انواع فرمون ها برای مدیریت کندو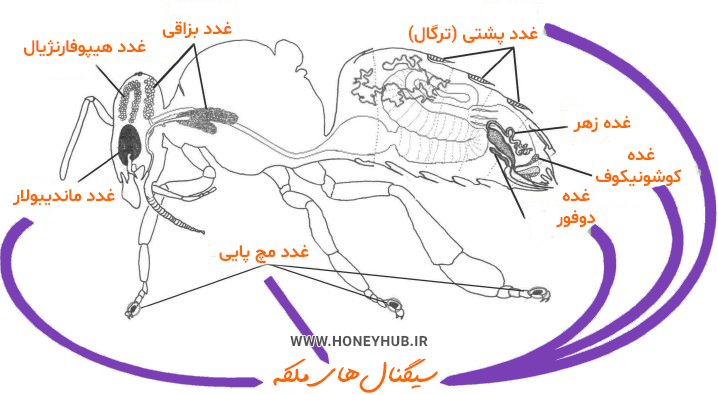 شکل 3- غدد ترشحی زنبور عسل
شکل 3- غدد ترشحی زنبور عسل شکل 4- آناتومی زنبور
شکل 4- آناتومی زنبور












Последние комментарии